肌原纤维形成过程中的细胞和分子事件
心脏是胚胎中最早分化和发挥功能的器官. 心肌细胞合成肌合成蛋白,这些蛋白组装成肌原纤维. 这些早期的肌原纤维负责第一次收缩和, 晚些时候, 生长中的心脏不断收缩,有丝分裂和细胞分裂. 细胞分裂后, 肌原纤维可以分解成它们的组成纤维和分子, 其中一些用于形成细胞分裂中使用的可收缩分裂沟(看到 研究兴趣).
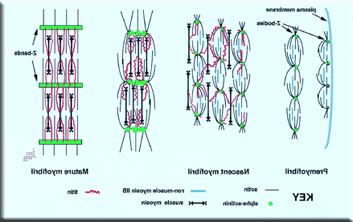
肌原纤维形成的确切发育级联尚不清楚. 随着细胞表面的扩展边缘的推进, 先前形成的肌原纤维被留在原位, 这将允许其他大的蛋白质(e.g.(肌肉肌球蛋白II和泰坦)以进入微角. 肌原纤维并不像有些人认为的那样,是成熟肌原纤维的分解产物. 肌原纤维前的z小体似乎在侧面相互融合,形成成熟肌原纤维的z带. 随着时间的推移, 成熟的肌原纤维和z带出现在z体线性排列的区域. 提出了肌原纤维形成的三个阶段模型,从肌原纤维前期到新生肌原纤维再到成熟肌原纤维. 肌原纤维位于心肌细胞的伸展边缘,由微原纤维组成. The sarcomeric equivalent of the Z-band in the premyofibril is a Z-body; both contain the cytoskeletal protein alpha actinin (100 kDA). 这种蛋白质主要存在于应力纤维中, 在非肌肉应力纤维和肌原纤维中,肌肉的z体和z带标志着肌体组织的边界. 我们的结果表明,肌原纤维在空间和时间上先于成熟的肌原纤维. 这些珠状的z型体附着在细胞表面,负责将肌动蛋白短丝附着在细胞表面. 最重要的是, 肌原纤维前含有非肌球蛋白IIB, 谁被认为是造成这些微型细胞中肌动蛋白丝的反极性排列的原因.
Rhee等. 证据表明,肌动蛋白丝捕获肌肉粗丝,并在z小体上新增肌肽分子,肌原纤维转化为新生肌原纤维. 相邻的前肌原纤维融合形成新生肌原纤维发生在z小体水平,并以至少两种蛋白质的结合为标志, Titin和zeugatin. 我们已经证明了胞浆蛋白, 在新生肌原纤维的z小体融合区可以检测到哪些, titin分子的一部分嵌入在z波段吗. 我们认为,广泛报道的肥厚心脏中心肌细胞重新启动胎儿肌纤维蛋白合成的能力可能与肌原纤维形成的胚胎程序的重新启动有关, 这是肌原纤维前模型.
在肌原纤维向肌原纤维过渡的过程中, 假设存在非肌球蛋白IIB丝与肌球蛋白II丝的交换,并且z小体生长并融合成z带. z小体最初表现为沿着肌原纤维前的α肌动蛋白的离散聚集体. 随着肌原纤维宽度的增加, z带似乎由横向排列的z体和连续的α肌动蛋白带组成. 肌原纤维形成的第二种模型提出,在扩张的心肌细胞周围形成的第一批原纤维作为肌原纤维聚集的临时支架. 最后, 还有第三种假设,即肌动蛋白丝和z带的复合物在空间上是分开的, IZI刷, 肌凝蛋白粗丝组彼此独立地组装在一起,由肌凝蛋白丝拼接在一起,然后插入到完全形成的肌原纤维的末端.
对肌原纤维形成过程中单细胞中的肌合成蛋白进行延时观察,可以对这些模型的各个方面进行测试. 我们实验室使用绿色荧光蛋白(GFP)长时间观察活细胞中的各种肌肉蛋白. 我们已经生成了许多细胞骨架蛋白的GFP融合蛋白,并观察到它们在扩散的胚胎鸡心肌细胞中遵循肌纤维形成的复杂调控.